







自噬(Autophagy)及其研究方法概述
一、背景
概念:目前根据发生过程分为三类:Macroautophagy,Microautophagy和Chaperone-mediated autophagy CMA), 大自噬(Macroautophagy)即我们说的自噬(autophagy);微自噬(Microautophagy):是指溶酶体主动、直接吞噬胞浆成分的一种方式;分子伴侣介导的自噬 (Chaperone-mediated autophagy,CMA):一些分子伴侣,如hsp70,能帮助未折叠蛋白转位入溶酶体。通常说的自噬泛指Macroautophagy.
自噬是细胞内的一种“自食(Self-eating)”的现象,凋亡是“自杀(Self-killing)”的现象,二者共用相同的刺激因素和调节蛋白,但是诱发阈值和门槛不同,如何转换和协调目前还不清楚. 自噬是指膜(目前来源还有争议,大部分表现为双层膜,有时多层或单层)包裹部分胞质和细胞内需降解的细胞器、蛋白质等形成自噬体(autophagosome),最后与溶酶体融合形成自噬溶酶体(autophagolysosome),降解其所包裹的内容物,以实现细胞稳态和细胞器的更新。自噬的步骤可以大概总结为下面四步:
步骤1:细胞接受自噬诱导信号后,在胞浆的某处形成一个小的类似“脂质体”样的膜结构,然后不断扩张,但它并不呈球形,而是扁平的,就像一个由2层脂双层组成的碗,可在电镜下观察到,被称为Phagophore,是自噬发生的铁证之一。
步骤2:Phagophore不断延伸,将胞浆中的任何成分,包括细胞器,全部揽入“碗”中,然后“收口”,成为密闭的球状的autophagosome,即“自噬体”。电镜下观察到自噬体是自噬发生的铁证之二。有2个特征:一是双层膜,二是内含胞浆成分,如线粒体、内质网碎片等。
步骤3:自噬体形成后,可与细胞内吞的吞噬泡、吞饮泡和内体融合(这种情况不是必然要发生的)。
步骤4:自噬体与溶酶体融合形成autolysosome,期间自噬体的内膜被溶酶体酶降解,2者的内容物合为一体,自噬体中的“货物”也被降解,产物(氨基酸、脂肪酸等)被输送到胞浆中,供细胞重新利用,而残渣或被排出细胞外或滞留在胞浆中。
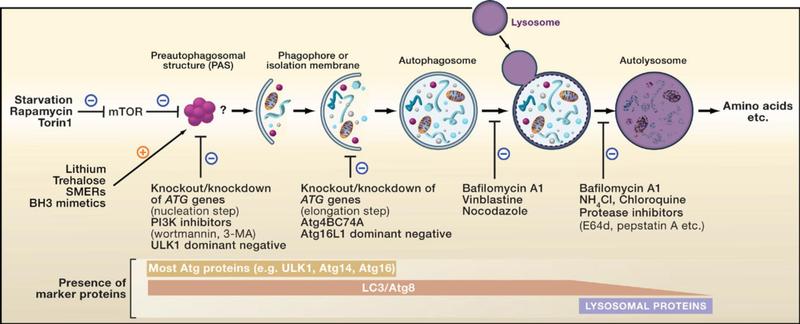
自噬的特性:
1)自噬是细胞消化掉自身的一部分,即self-eating,初一看似乎对细胞不利。事实上,细胞正常情况下很少发生自噬,除非有诱发因素的存在。这些诱发因素很多,也是研究的热门。既有来自于细胞外的(如外界中的营养成分、缺血缺氧、生长因子的浓度等),也有细胞内的(代谢压力、衰老或破损的细胞器、折叠错误或聚集的蛋白质等)。由于这些因素的经常性存在,因此,细胞保持了一种很低的、基础的自噬活性以维持自稳。
2)自噬过程很快,被诱导后8min即可观察到自噬体(autophagosome)形成,2h后自噬溶酶体(autolysosome)基本降解消失。这有利于细胞快速适应恶劣环境。
3)自噬的可诱导特性:表现在2个方面,第一是自噬相关蛋白的快速合成,这是准备阶段。第二是自噬体的快速大量形成,这是执行阶段。
4)批量降解:这是与蛋白酶体降解途径的显著区别
5)“捕获”胞浆成分的非特异性:由于自噬的速度要快、量要大,因此特异性不是首先考虑的,这与自噬的应急特性是相适应的。
6)自噬的保守性:由于自噬有利于细胞的存活,因此无论是物种间、还是各细胞类型之间(包括肿瘤细胞),自噬都普遍被保留下来
自噬过程的调控:
从上面总结的自噬特点中可以看出,自噬这一过程一旦启动,必须在度过危机后适时停止,否则,其非特异性捕获胞浆成分的特性将导致细胞发生不可逆的损伤。这也提醒我们在研究自噬时一定要动态观察,任何横断面的研究结果都不足以评价自噬的活性。目前,已经报告了很多因素能诱导细胞发生自噬,如饥饿、生长因子缺乏、微生物感染、细胞器损伤、蛋白质折叠错误或聚集、DNA损伤、放疗、化疗等等,这么多刺激信号如何传递的、哪些自噬蛋白接受信号、又有哪些自噬蛋白去执行等很多问题都还在等待进一步解答中。
关于传递自噬信号的通路目前比较肯定的有:
抑制类
1)Class I PI3K pathway (PI-phosphatidylinositol,磷脂酰肌醇)与IRS (Insulin receptor substrate) 结合,接受胰岛素受体传来的信号(血糖水平高抑制自噬)
2)mTOR pathway(mammalian target of rapamycin)
mTOR在人类中的同源基因是FRAP1(FK506 binding protein 12-rapamycin associated protein 1),是一个丝/苏氨酸蛋白激酶。能接受多种上游信号,如Class I PI3K、IGF-1/2、MAPK,能感受营养和能量的变化,rapamycin是最典型最常用的自噬激动剂.
激活类
1)Class III PI3K
结构上类似于Class I PI3K,但作用相反。3-MA是Class III PI3K的抑制剂,因此3-MA可以作为自噬的抑制剂.
二、自噬的研究方法:
汉恒生物已开发并完善了多套自噬相关的研究体系和工具,并积累了丰富的自噬研究经验。
正常培养的细胞自噬活性很低,不适于观察,因此,必须对自噬进行人工干预和调节,经报道的工具药有:
(一)自噬诱导剂
1)Bredeldin A / Thapsigargin / Tunicamycin :模拟内质网应激
2)Carbamazepine/ L-690,330/ Lithium Chloride(氯化锂):IMPase 抑制剂(即Inositol monophosphatase,肌醇单磷酸酶)
3)Earle's平衡盐溶液:制造饥饿
4)N-Acetyl-D-sphingosine(C2-ceramide):Class I PI3K Pathway抑制剂
5)Rapamycin:mTOR抑制剂(这是最常用的)
6)Xestospongin B/C:IP3R阻滞剂
(二)自噬抑制剂
1)3-Methyladenine(3-MA):(Class III PI3K) hVps34 抑制剂
2)Bafilomycin A1:质子泵抑制剂
3)Hydroxychloroquine(羟氯喹)
除了选用上述工具药外,一般还需结合遗传学技术对自噬相关基因进行干预:包括反义RNA干扰技术(Knockdown)、突变株筛选、外源基因导入等。
细胞经诱导或抑制后,需对自噬过程进行观察和检测,常用的策略和技术有:
1)观察自噬体的形成
由于自噬体属于亚细胞结构,普通光镜下看不到,因此,直接观察自噬体需在透射电镜下。Phagophore的特征为:新月状或杯状,双层或多层膜,有包绕胞浆成分的趋势。自噬体(AV1)的特征为:双层或多层膜的液泡状结构,内含胞浆成分,如线粒体、内质网、核糖体等。自噬溶酶体(AV2)的特征为:单层膜,胞浆成分已降解。(autophagic vacuole,AV)
2)在荧光显微镜下采用GFP-LC3等融合蛋白来示踪自噬形成:(常用)
GFP-LC3单荧光指示体系:由于电镜耗时长,不利于监测(Monitoring)自噬形成。我们利用LC3在自噬形成过程中发生聚集的现象开发出了GFP-LC3指示技术:无自噬时,GFP-LC3融合蛋白弥散在胞浆中;自噬形成时,GFP-LC3融合蛋白转位至自噬体膜,在荧光显微镜下形成多个明亮的绿色荧光斑点,一个斑点相当于一个自噬体,可以通过计数来评价自噬活性的高低。汉恒生物已开发出高效的评价用GFP-LC3病毒载体,通过瞬时高效感染细胞,配合活细胞工作站成功评价自噬流。
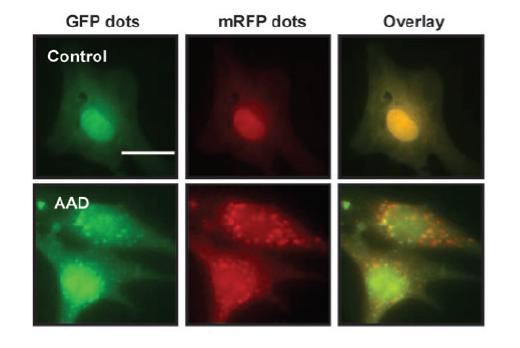
双荧光指示体系:汉恒生物科技(上海)有限公司已开发出用于表达mRFP-GFP-LC3融合蛋白的病毒产品。mRFP用于标记及追踪LC3,GFP的减弱可指示溶酶体与自噬小体的融合形成自噬溶酶体,即由于GFP荧光蛋白对酸性敏感,当自噬体与溶酶体融合后GFP荧光发生淬灭,此时只能检测到红色荧光。

我们在显微镜成像后红绿荧光merge后通过merge后出现的黄色斑点即只是自噬体.红色的斑点指示自噬溶酶体,通过不同颜色斑点的计数可以清晰的看出自噬流的强弱。
如下图:细胞转染mRFP-GFP-LC3病毒后给予氨基酸剥夺处理2小时后出现明显增强的自噬以及自噬流。

3)利用Western Blot检测LC3-II/I比值的变化以评价自噬形成(LC3抗体购自sigma,L8918)。
自噬形成时,胞浆型LC3(即LC3-I)会酶解掉一小段多肽,转变为(自噬体)膜型(即LC3-II),因此,LC3-II/I比值的大小可估计
自噬水平的高低。
(Note:LC3抗体对LC3-II有更高的亲和力,会造成假阳性。方法2和3需结合使用,同时需考虑溶酶体活性的影响。)
4) 利用Western Blot检测p62蛋白来评价自噬以及自噬流的强弱:起初自噬所降解的底物被认为是随机的,但是后来的研究表明有些蛋白是seletively降解的,在这些蛋白之中研究的最为透彻的是蛋白p62, p62 is selectively incorporated into autophagosomes through direct binding to LC3 and is efficiently degraded by autophagy ; thus, the total cellular expression levels of p62 inversely correlate with autophagic activity. (p62蛋白水平的多少与自噬流的强弱有着反比例关系)
三、自噬与肿瘤的关系:
与凋亡(在肿瘤细胞中一般都存在缺限)不同,自噬是被优先保留的。无论是肿瘤细胞还是正常细胞,保持一种基础、低水平的自噬活性是至关重要的。因为细胞中随时产生的“垃圾”(破损或衰老的细胞器、长寿命蛋白质、错误合成或折叠错误的蛋白质等等)都需要及时清除,而这主要靠自噬来完成,因此,自噬具有维持细胞自稳的功能;如果将自噬相关基因突变失活,如神经元会发生大量聚集蛋白,并出现神经元退化。同时,自噬的产物,如氨基酸、脂肪酸等小分子物质又可为细胞提供一定的能量和合成底物,可以说,自噬就是一个“备用仓库”。如Atg-5缺陷的小鼠在出生后喝上第一口奶之前就会饿死。更重要的是,自噬活性可在代谢应激(饥饿、生长因子缺乏、射线、化疗等)时大大增强,表现为胞浆中迅速涌现大量自噬体,有利于细胞的存活。
鉴于自噬的上述作用,自噬可为肿瘤细胞带来几大好处:
1)肿瘤细胞本身就具有高代谢的特点,对营养和能量的需求比正常细胞更高,但肿瘤微环境往往不能如意,如肿瘤发生初始期到血管发生之前、肿瘤长大发生血管崩塌时、肿瘤细胞脱离原发灶游走时等都会出现营养不足或供应中断,而此时提高自噬活性可以有助于度过这一危机。
2)当化疗、放疗后,肿瘤细胞会产生大量的破损细胞器、损坏的蛋白质等有害成分,而此时提高自噬活性可及时清除这些有害物质,并提供应急的底物和能量为修复受损DNA赢得时间和条件.由于自噬减少了肿瘤细胞在代谢应激时发生坏死的机会,而对于肿瘤细胞群体而言,需要一部分细胞发生坏死,以引发适度的炎症(有利于血管的长入、吸引免疫细胞分泌生长因子等)。研究发现,很多类型的肿瘤在代谢应激时会“组成性”活化PI3K信号以抑制自噬(由于凋亡通路已受阻,抑制自噬会促进坏死),但具体机制尚不清楚。
自噬与肿瘤的关系可能是双重的。①对不同的细胞,自噬的作用可能不同。②相同的细胞在不同的外部因素作用时,自噬的作用可能不同。③在肿瘤发生发展的不同阶段,自噬的作用可能不同。肿瘤生长的早期阶段自噬增强,是由于此时肿瘤的血管化作用不足,癌细胞的营养供给有限,需要通过自噬为自身提供营养。肿瘤进入发展阶段后基因变异积累,使包括 Beclin 1在内的众多抑癌基因失活,自噬活力降低。④对单个细胞和对整个肿瘤阻滞的作用可能不同。自噬功能不全的细胞易于坏死,但是坏死组织产生的细胞因子(包括部分生长因子)反而会促进肿瘤的生长。上述各种假设均有待证实。肿瘤为细胞分化障碍性的疾病已得到肯定,但自噬在肿瘤细胞的分化抑制过程中起着什么样的作用,自噬水平提高是抑制分化甚至导致去分化还是促进分化等问题尚未解决。
- 浏览 74952 次

